

La neurofisiología elemental trata de estudiar el
comportamiento de neuronas o grupos de neuronas aisladas. Los
hechos establecidos por la neurofisiología elemental
pueden ser aprovechados por la teoría matemática de
redes neuronales para construir modelos matemáticos que
permitan identificar fenómenos neurofisiológicos
como la memoria aprendizaje. y el
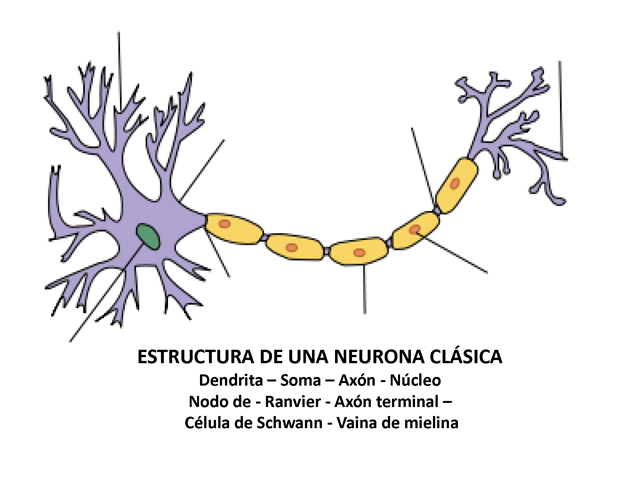
ESTRUCTURA DE UNA NEURONA CLÁSICA Dendrita – Soma
– Axón - Núcleo Nodo de - Ranvier -
Axón terminal – Célula de Schwann - Vaina de
mielina

Los principales hechos establecidos por la neurofisiología
elemental tenidos en cuenta en la construcción de modelos
de redes neuronales son: Un cerebro gran cantidad de neuronas. El
número de neuronas de un cerebro humano se ha estimado en
más de 1011 neuronas. Las neuronas consisten en un cuerpo
celular, una estructura dendrítica arbórea y un
axón. Las neuronas son células vivas con un
metabolismo similar al encontrado en el resto de células.
Así el cuerpo celular o soma contiene un núcleo,
vesículas, mitocondrias y otros orgánulos. A
diferencias de otras células, además posee
dendritas y axón. Las dendritas forman una estructura
arbórea inmensa que puede extenderse por amplias
áreas de un cerebro, los axiones pueden llegar a tener
más de un metro de longitud. Las neuronas generan
potenciales eléctricos. Los potenciales eléctricos
o potenciales de acción, también llamados pulsos
eléctricos o chispas de voltaje, son fenómenos
electrofisiológicos provocados porque las membranas
celulares de las neuronas tienen propiedades activas que las
hacen excitables o sensibles a potenciales eléctricos
procedentes de otras neuronas. Estos potenciales
eléctricos se originan usualmente en el extremo del
axón y se propagan a lo largo de su longitud. Los
potenciales eléctricos son los mecanismos básicos
para la comunicación entre neuronas. Los potenciales de
acción pueden considerarse como señales
eléctricas que una neurona envía a otras. Cada
neurona recibe muchas señales procedentes de otras
neuronas (potencial convergente) y a su vez envía
señalas a muchas otras (potencial emergente).

Las neuronas están funcionalmente polarizadas. Esto es,
las neuronas reciben señales eléctricas a
través de sus dendritas, procesan y superponen dichas
señales en el soma y envían una respuesta a otras
neuronas a través de su axón. La unión entre
el axón de una neurona y las dendritas de otra neurona se
llama sinapsis. Las sinapsis pueden ser eléctricas o
químicas. Una sinapsis química está formada
por un emisor presináptico y un receptor
postsináptico que están separadas por un espacio
sináptico. Cuando un impulso llega al final de un
axón, se dispara una cadena de reacciones químicas
fisiológicas en la presinapsis, que conllevan la
liberación de sustancias químicas en el espacio
sináptico. Las substancias liberadas se denominan
neurotransmisores. Estos se difunden pasivamente a lo largo del
espacio sináptico produciendo cambios en el potencial de
la membrana postsináptica. El Principio de Dale, establece
que una neurona es o bien excitatoria o bien inhibitoria. Es
excitatoria si el potencial de la membrana postsináptica
se incrementa, hecho conocido como "despolarización".
Cuando una neurona se despolariza se facilita la
generación de un potencial de acción en la neurona
postsináptica. Si por el contrario el potencial decrece la
neurona es inhibitoria. La hiperpolarización que puede
llegar a sufrir una neurona inhibitoria impide la
generación de potencial de acción

NEUROFISIOLOGÍA FUNDAMENTOS BÁSICOS Y POTENCIALES
DE ACCIÓN. • La neurofisiología tiene como
objetivo comprender el funcionamiento del sistema nervioso, y el
buen funcionamiento del sistema nervioso depende de que el flujo
de información que este se encarga de transmitir, sea
rápida y eficiente entre las neuronas y sus efectores.
• La información se transmite utilizando
señales eléctricas, que se propagan a lo largo de
los axones de las neuronas. Esta señal eléctrica se
conoce como impulso nervioso, o potencial de acción.

El conocimiento de estas señales eléctricas se debe
al al Dr. Luigi Galvani, en el siglo XVIII, que demostró
conectando unos electrodos a la médula espinal de una rana
(muerta obviamente), que los tejidos animales son sensibles a la
corriente eléctrica, esta corriente eléctrica
depende de las propiedades eléctricas de las membranas,
que juegan un papel de suma importancia en este sentido. El
término clave en este ámbito es el potencial de
membrana, que es la diferencia de potencial electroquímico
a ambos lados de una membrana que separa dos soluciones de
diferente concentración de iones. Cuando las neuronas no
generan impulsos, se dice que presentan un potencial de membrana
en reposo, y que es así si nada lo altera.

En reposo, el potencial de membrana de las neuronas es
electronegativo, rondando la cifra de -70 mV. Tal como
decía la definición que encontramos en el
párrafo anterior, los movimientos iónicos son por
gradiente químico (de concentración) y
eléctrico (diferencia de cargas positivas vs negativas). Y
aquí comenzamos con las leyes y fórmulas que tanto
nos gustan, la Ley de Ohm, que relaciona el potencial (diferencia
de potencial) (V), con la conductancia (g), con la cantidad de
corriente que fluirá (I) , mediante esta fórmula:
I=gV. Lo primero que podemos observar es que tanto si la
conductancia, o la diferencia de potencial, es cero, no
habrá movimiento iónico. Mencionamos aquí el
equilibrio de Gibbs – Donnan que se produce entre los iones
que pueden atravesar la membrana y los que no son capaces de
hacerlo. Las composiciones en el equilibrio se ven determinadas
tanto por las concentraciones de los iones como por sus cargas.
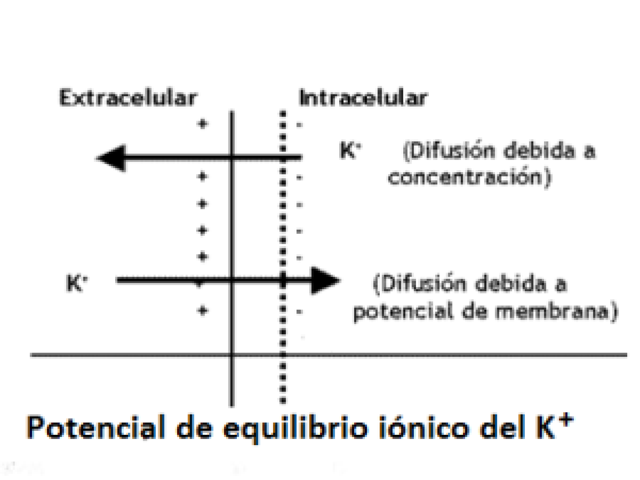
En el potencial de equilibrio, el movimiento NETO del ion es
nulo, eso no quiere decir que no haya movimiento, si no que la
suma de movimientos se da de forma neutra, es un equilibrio
dinámico. Se produce cuando se equilibran las
concentraciones con las cargas. como ejemplifica la siguiente
imagen:

Y ahora entra en juego la ecuación de Nerst, que da el
valor (en mV) de potencial necesario para que un ión
esté en equilibrio. Esta ecuación, adaptada a
37ºC, y en la forma más práctica, es la
siguiente: Vinterior – Vexterior = 61,4 · log
([Exterior] /[interior]) El potencial de membrana en reposo
depende del movimiento pasivo de iones, pero como indica el
equilibrio de Gibbs – Donnan, la membrana tiene unas
características de permeabilidad muy diferentes para cada
ion, la ecuación de Goldman tiene en cuenta la
permeabilidad, a 37ºC, es una mejora de la de Nerst: FEM =
Vinterior – Vexterior. C = Concentración. P:
Permeabilidad de la membrana al ión.
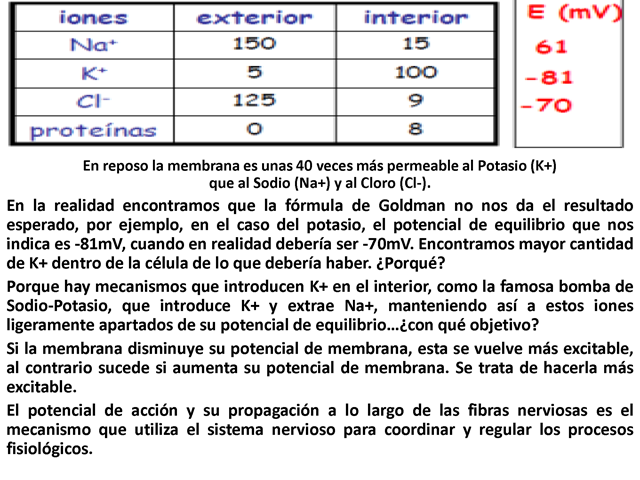
En reposo la membrana es unas 40 veces más permeable al
Potasio (K+) que al Sodio (Na+) y al Cloro (Cl-). En la realidad
encontramos que la fórmula de Goldman no nos da el
resultado esperado, por ejemplo, en el caso del potasio, el
potencial de equilibrio que nos indica es -81mV, cuando en
realidad debería ser -70mV. Encontramos mayor cantidad de
K+ dentro de la célula de lo que debería haber.
¿Porqué? Porque hay mecanismos que introducen K+ en
el interior, como la famosa bomba de Sodio-Potasio, que introduce
K+ y extrae Na+, manteniendo así a estos iones ligeramente
apartados de su potencial de equilibrio…¿con
qué objetivo? Si la membrana disminuye su potencial de
membrana, esta se vuelve más excitable, al contrario
sucede si aumenta su potencial de membrana. Se trata de hacerla
más excitable. El potencial de acción y su
propagación a lo largo de las fibras nerviosas es el
mecanismo que utiliza el sistema nervioso para coordinar y
regular los procesos fisiológicos.

Un potencial de acción o también llamado impulso
eléctrico, es una onda de descarga eléctrica que
viaja a lo largo de la membrana celular modificando su
distribución de carga eléctrica. Los tejidos
nervioso y muscular son excitables, esta es la base para nuestro
movimiento muscular. Un estimulo eléctrico debe superar un
umbral, es decir, debe despolarizar lo suficiente la membrana
como para poder generar un potencial de acción. Este
potencial umbral varía, pero normalmente está en
torno a -55 a -50 milivoltios sobre el potencial de reposo de la
célula, lo que implica que la corriente de entrada de
iones sodio supera la corriente de salida de iones potasio. El
potencial de acción puede explicarse en base a cambios en
la conductancia de la membrana hacia ciertos iones, jugando un
papel importante los canales iónicos, y más en este
sentido, los dependientes de voltaje. Propiedades de los canales
de Na+ sensibles al voltaje. Cuando se abre, el Na+ pasa del
exterior al interior celular, lo que produce una
despolarización. Cuando más se despolariza la
membrana, más canales se abren, mayor
despolarización, ejemplo claro de retroalimentación
positiva, un estimulo inicial refuerza que se siga fortaleciendo
ese estimulo. La membrana tiende a alcanzar el potencial de
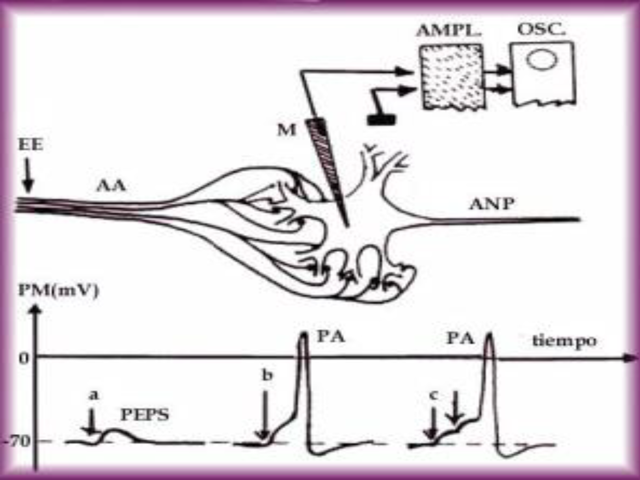
equilibrio del Na+ = 55mV. Fases de un Potencial de Acción
Todo potencial de acción se rige por unas fases, como
muestra la siguiente imagen.
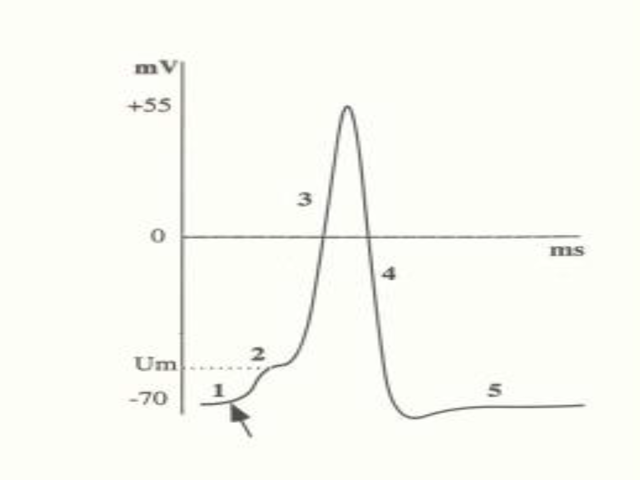

1- Nivel de reposo. 2- Despolarización. 3- Potencial
invertido. 4- Repolarización. 5-
Posthiperpolarización. Obsérvese la fase
refractaria número 4, donde el valor se acerca más
a EK de lo que estaba en reposo. Dos periodos refractarios a
destacar, que determinan la frecuencia de los potenciales de
acción y hace que estos sean UNIDIRECCIONALES
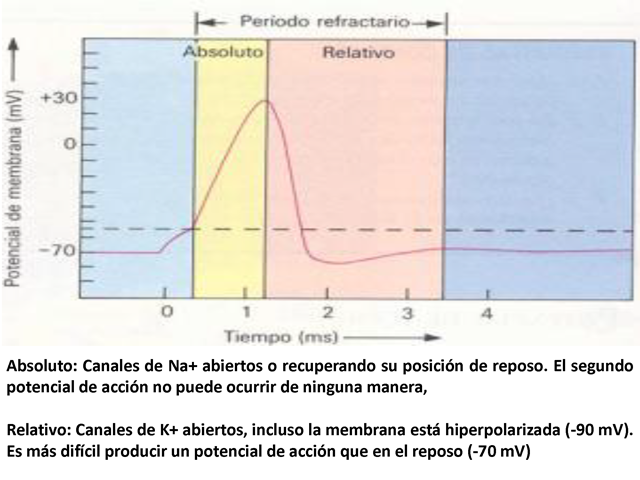
Absoluto: Canales de Na+ abiertos o recuperando su
posición de reposo. El segundo potencial de acción
no puede ocurrir de ninguna manera, Relativo: Canales de K+
abiertos, incluso la membrana está hiperpolarizada (-90
mV). Es más difícil producir un potencial de
acción que en el reposo (-70 mV)
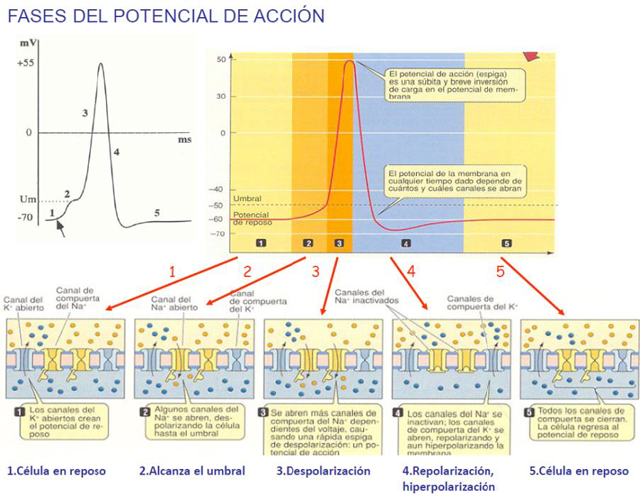

LEY DEL “TODO O NADA” Los PA tienen amplitud
constante, no hay PA ni grandes ni pequeños, o hay, o no,
en una misma neurona. La amplitud puede variar según la
neurona. Siempre van desde -70 mV hasta el potencial de
equilibrio del Na+, 55 mV, punto que se tiende a alcanzar.
Duración de los potenciales de acción En total,
incluyendo periodo refractario absoluto, aprox 1 ms. Ergo,
delimitado por lo anterior, frecuencia máxima de PA en una
fibra es de 1000 PA/s. Determinada por la fuerza de las
señales, cuanto más intensas, mayor frecuencia de
PA. La conducción de un Potencial de Acción La
estructura de las neuronas está especialmente adaptada
para la comunicación de estos impulsos nerviosos, el PA se
genera en el segmento inicial de la neurona, y se propaga a lo
largo de esta, de extremo a extremo. Esta conducción es
autopropagante.

CONDUCCIÓN DEL POTENCIAL DE ACCIÓN EN UN
AXÓN NO MIELINICO La llegada de un PA invierte la
polaridad de la membrana Las cargas de diferente signo son
atraídas por las de regiones vecinas La región
siguiente se despolariza parcialmente y se produce la apertura de
canales de Na+ regulados por el voltaje… y se produce un
PA. La región donde comenzó el PA se vuelve
refractaria momentáneamente por lo que la
propagación tiene lugar en una sola dirección.
CONDUCCIÓN DEL PA EN UN AXÓN MIELÍNICO
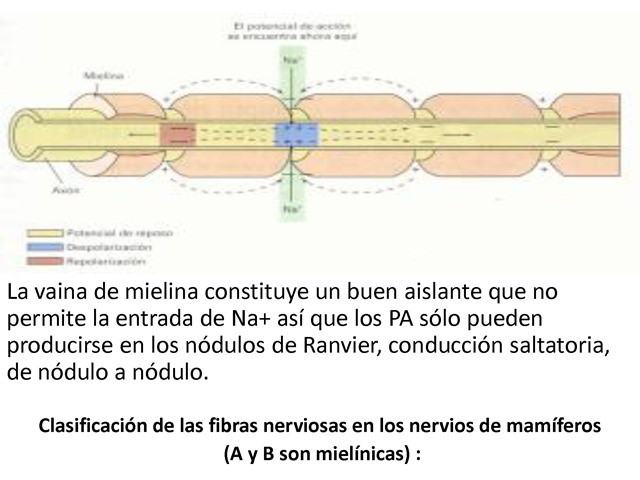
La vaina de mielina constituye un buen aislante que no permite la
entrada de Na+ así que los PA sólo pueden
producirse en los nódulos de Ranvier, conducción
saltatoria, de nódulo a nódulo.
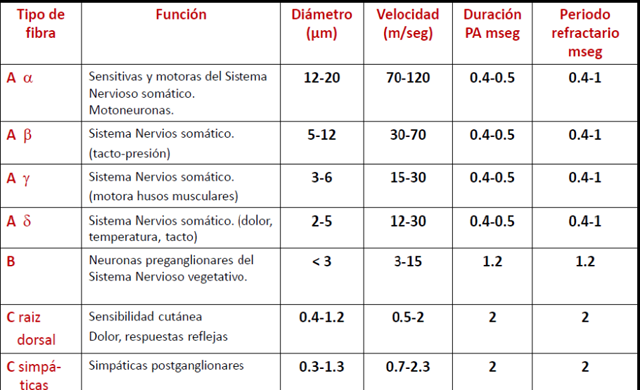
Clasificación de las fibras nerviosas en los nervios de
mamíferos (A y B son mielínicas) :


La neurofisiología es una rama de la neurología que
estudia al sistema nervioso desde el punto de vista funcional,
generalmente valora el componente eléctrico. El
profesional que hace estas valoraciones se llama
Neurofisiólogo quien es un neurólogo con
especialización en la realización de estudios
funcionales. Si el paciente es un niño o un adolescente lo
ideal es que el profesional tratante sea un Neurofisiólogo
Pediatra.
Trabajos relacionados
Ver mas trabajos de Anatomia |
Nota al lector: es posible que esta página
no contenga todos los componentes del trabajo original (pies de página,
avanzadas formulas matemáticas, esquemas o tablas complejas, etc.).
Recuerde que para ver el trabajo en su versión original completa, puede
descargarlo desde el menú superior.
no contenga todos los componentes del trabajo original (pies de página,
avanzadas formulas matemáticas, esquemas o tablas complejas, etc.).
Recuerde que para ver el trabajo en su versión original completa, puede
descargarlo desde el menú superior.
Todos los documentos disponibles en este
sitio expresan los puntos de vista de sus respectivos autores y no de
Monografias.com. El objetivo de Monografias.com es poner el conocimiento
a disposición de toda su comunidad. Queda bajo la responsabilidad de
cada lector el eventual uso que se le de a esta información. Asimismo,
es obligatoria la cita del autor del contenido y de Monografias.com como
fuentes de información.
sitio expresan los puntos de vista de sus respectivos autores y no de
Monografias.com. El objetivo de Monografias.com es poner el conocimiento
a disposición de toda su comunidad. Queda bajo la responsabilidad de
cada lector el eventual uso que se le de a esta información. Asimismo,
es obligatoria la cita del autor del contenido y de Monografias.com como
fuentes de información.
No hay comentarios:
Publicar un comentario